
Приглядитесь пристально к этому чудному пейзажу, и вы поймете, что все его участники очень похожи. На первый взгляд – не очень. Ну что может быть общего у раскидистого дерева, жующей его листья гусеницы, растущих из ствола грибочков, устилающей землю травы и благородного оленя, отдыхающего на ней в тени дерева? И тем не менее на микроскопическом уровне они все удивительно похожи: в их клетках есть ядро, набитое ДНК и отделенное от цитоплазмы двумя мембранами, целая система мембранных полостей и пузырьков в самой цитоплазме, ниточки и трубочки цитоскелета, а еще – миловидные митохондрии, которые снабжают клетку энергией. Все эти структуры присутствуют почти во всех клетках любого животного, гриба, растения, как и у любого одноклеточного протиста – у всех тех, кого биологи называют эукариотами.
В отличие от них, у так называемых прокариот – бактерий и архей – нет ни ядра, ни эндоплазматической сети, ни митохондрий, они представляют собой мелкие (намного меньше средней эукариотной клетки) одноклеточные организмы, устроенные относительно просто, хотя не стоит думать, что они такие уж простачки. Прокариоты – поразительные машины выживания. Они обитают повсюду – от глубочайших океанских глубин до высочайших атмосферных высот. Они способны обмениваться генами друг с другом напрямую в процессе горизонтального переноса и благодаря этому эволюционируют быстрыми темпами, адаптируясь к всевозможным экстремальным условиям и приобретая всякие полезные сверхспособности. Они даже общаются между собой и посредством чувства кворума объединяются в биопленки, которые агрессивно колонизируют доступные поверхности, включая ваши зубы и стенки внутренних органов. Они жили на Земле задолго до появления эукариот и, возможно, еще их переживут.
И все же, при всех своих удивительных свойствах, без характерной для эукариот архитектуры прокариоты обречены вечно быть малютками и никогда не познают истинной многоклеточности. Прокариоты могут хоть всю планету покрыть глобальной биопленкой, но лугами, лесами, подвижными телами и грандиозными сооружениями ее заполонили именно эукариоты.
Переход от «классической» прокариотической модели жизни к «модерновой» эукариотической был одним из важнейших событий в истории Земли. Причем событием, вероятно, уникальным. Некоторые сложные структуры появлялись в процессе эволюции неоднократно и независимо у разных групп живых организмов – многоклеточные формы, глаза, покровы, крылья. Но эукариотная клетка за три-четыре миллиарда лет земной жизни возникла лишь единожды. По крайней мере все ныне живущие эукариоты являются потомками одной-единственной клетки-прабабули, возникшей в результате фантастического слияния двух прокариот, синергия которых позволила ей эволюционировать в совершенно новых направлениях, недостижимых ни для одной отдельно взятой прокариоты.
Мир, каким мы его видим (и тот факт, что мы его в принципе видим, ведь глаза – это эукариотическая фишка), есть результат счастливого союза, столь невероятного, что его могло бы и не случиться вовсе. И тогда планета по-прежнему была бы заселена исключительно прокариотическими микробами, и не было бы в мире ни тебя, ни меня, ни оленя, ни коня, ни опят у того пня, ни луня, ни ясеня.
Краткая история симбиогенеза
В 1883 году немецкий ботаник Андреас Шимпер поделился своими наблюдениями за делением хлоропластов – клеточных органелл, в которых происходит фотосинтез (они имеются у растений и разных протистов): по его мнению, оно было очень похоже на деление свободноживущих цианобактерий. На основании этого ученый высказал осторожную мысль, что растения появились в результате симбиоза. В 1905 году его русский коллега Константин Мережковский сформулировал теорию, согласно которой хлоропласты некогда были цианобактериями, которые поселились в более крупных клетках, став их эндосимбионтами. Он назвал это симбиогенезом. В 1922 году эксцентричный американский биолог Айван Уоллин по прозвищу Человек-Митохондрия заявил о симбиогенетической природе митохондрий. Но в первой половине XX века теория симбиогенеза не находила поддержки у научного сообщества и если не критиковалась жестко, то попросту игнорировалась.

Все изменилось после 1967 года, когда в «Журнале теоретической биологии» вышла неординарная статья выдающейся американской исследовательницы Линн Маргулис (статья опубликована под фамилией Саган, которая досталась Линн от бывшего мужа, Карла Сагана, ставшего впоследствии знаменитым популяризатором астрономии). В ней Маргулис провозгласила, что митохондрии и хлоропласты – это бывшие бактерии, в древности проглоченные другими микробами: вот почему эти органеллы до сих пор сохраняют свои собственные крошечные геномы и внешне похожи на бактерий. Блестящая статья, вобравшая в себя аргументы из цитологии, биохимии, генетики, микробиологии, экологии, геологии и палеонтологии, бросала вызов всеобщему убеждению, что митохондрии возникли сами по себе внутри клеток в процессе эволюции. Тогда, полвека назад, слова «эндосимбиоз» и «симбиогенез» можно было лишь шептать про себя в уединенном месте в полной темноте. Издатели отказывали Маргулис раз пятнадцать, прежде чем напечатать ее работу.
Неудивительно, что она тут же подверглась беспощадной критике, на которую Маргулис, будучи бунтаркой, отвечала с неменьшей жесткостью. «Я не считаю свои теории спорными, – говорила она. – Я считаю их верными». Впоследствии филогенетические исследования доказали ее правоту. Сегодня мы знаем, что митохондриальная ДНК имеет много общего с ДНК свободноживущих бактерий. И даже догадываемся, с какими бактериями митохондрии состоят в наиближайшем родстве. Чаще всего ближайшими родственниками митохондрий называют пурпурных альфа-протеобактерий из группы Rickettsiales. Хотя прошлогодний анализ морских альфа-протеобактерий показал, что родство не такое уж и близкое: скорее митохондрии происходят от более древней линии протеобактерий, отщепившейся до того, как появился предок современных альф.
В 1977 году, через десять лет после статьи Линн Маргулис, произошла еще одна революция во взглядах на живую природу. Американский микробиолог Карл Вёзе изучал микробов-метаногенов, как вдруг внезапно понял, что бактериями они не являются. Это были первые известные науке представители архей – прокариот, которые, может, и похожи внешне на бактерий, но биохимически сильно отличаются от них. До того момента биологии были известны две крупные группы живых организмов – бактерии и эукариоты. Отныне же их было три. Как будто все разглядывали глобус с северным и южным полушариями, а Вёзе, вежливо кхекнув, раздвинул их и указал на еще одно, доселе сокрытое. Что еще интересно, археи по ряду признаков оказались ближе к эукариотам, нежели к бактериям, будучи притом типичными прокариотами – без ядра и митохондрий.
Вёзе полагал, что археи и эукариоты – это сестринские ветви, которые разделились вскоре после отделения их общего ствола от бактериального, очень рано в истории клеточной жизни. Однако дальнейшие генетические исследования и открытия новых групп архей ясно дали понять, что ветвь эукариот скромно растет из пышной кроны архейного древа и, по сути, является специализированной разновидностью архей. Просто в какой-то момент эти продвинутые археи обзавелись ядром, характерным цитоскелетом, эндоплазматической сетью и бактериальными симбионтами – митохондриями. Так два ствола соприкоснулись ветвями, и они привились друг к другу и породили новую веточку, разросшуюся в целый ствол. Древо жизни замкнулось в кольцо, породившее новое, эукариотическое древо.
Из варяг в эукариоты
Несколько лет назад мы узнали, где именно на древе архей находится место, откуда есть пошли эукариоты, а заодно и познакомились с нашими ближайшими современными архейными родственниками.
Научной группе из Швеции и Норвегии удалось собрать неполные геномы трех архей из пробы грунта, поднятой с глубины 3283 метра в Северном Ледовитом океане, недалеко от гидротермальной зоны «Замок Локи» на склоне срединно-океанического хребта. В научной статье 2015 года этих микробов нарекли локиархеями. Они оказались близкими к эукариотам, как никто другой из архей. Даже ближе к эукариотам, чем ко всем другим известным археям. Некоторые из их генов кодируют белки, считавшиеся уникальными для эукариот, в том числе связанные с регуляцией фагоцитоза (захвата объектов из внешней среды) и пузырькового транспорта, а также белки «локиактины», весьма похожие на актины, из которых строятся цитоскелетные нити в эукариотных клетках.
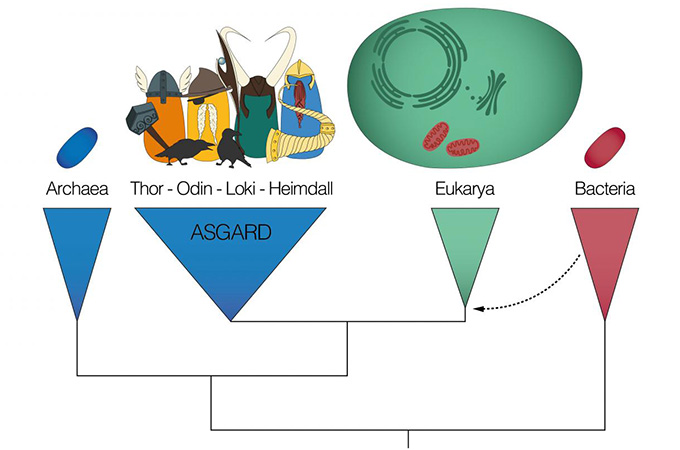
После этого знаменательного открытия ученые стали целенаправленно искать близких родственников локиархей – и нашли. В 2016 году американские специалисты представили миру торархей, чьи геномы были реконструированы по кусочкам ДНК из донных осадков эстуария реки Уайт-Оук на восточном побережье США. А в начале 2017 года международная научная группа, включившая первооткрывателей локи- и торархей, объявила об обнаружении одинархей в горячих источниках Йеллоустоуна и Новой Зеландии, о переименовании двух наиболее близких к эукариотам локиархей в хеймдалльархей с включением в них образца из залива Орхус в Дании, ну и о логичном объединении всей этой плеяды богоназванных архей в группу Асгард – в честь одноименного небесного города из скандинавской мифологии.
(Обновление. В 2019 году к ним добавились хельархеи, близкие к локиархеям и названные в честь Хель, дочери Локи, повелительницы мира мертвых. А в 2020-м – еще и гердархеи, близкие к хеймдалльархеям и названные в честь великанши Герд, богини плодородной почвы. Об этих двух группах можно почитать в нашем обзоре.)
Общий предок всех эукариот был одной из таких асгардархей – возможно, хеймдалльархеей или кем-то из ранних представителей группы. Он мог обладать уже весьма продвинутой анатомией – гибкой наружной мембраной, допускающей фагоцитоз, мембранными структурами внутри клетки, может быть, даже подобием ядра. Такие черты наряду с некоторыми физиологическими сходствами с эукариотами прогнозируются для асгардархей на основании анализа их генов и белковых последовательностей. Прежде чем приобрести бактериального симбионта, древние археи из Асгарда, похоже, уже двигались по магистральному пути эукариотизации.
Интриги добавляет тот факт, что самих микробов никто из открывших их ученых не видел воочию. Их гены были выявлены сами по себе – методами метагеномики: секвенируется ДНК сразу всех организмов в образце, затем гены и геномы собираются по фрагментам. В связи с этим подтвердить или опровергнуть прогнозы относительно морфологии и физиологии асгардархей пока не представляется возможным. Вырастить их в лаборатории очень сложно – они экстремофилы по натуре, обитают в условиях низкого содержания кислорода, метаболизм их крайне медленный, и некоторые из них, как предсказывают расчеты, делятся раз в десять лет! Мало кто из ученых вообще решается их культивировать.
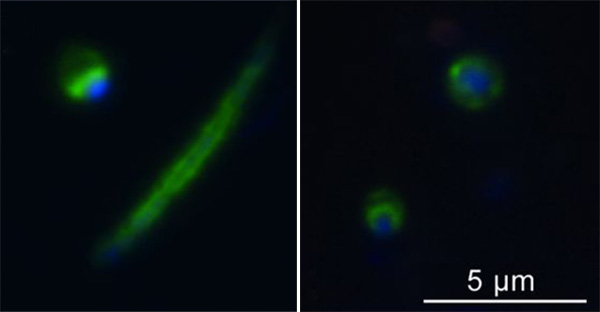
Меж тем независимая группа специалистов из Чехии и Румынии утверждает, что им удалось-таки увидеть хеймдалль- и локиархей в образцах донных осадков румынских озер с помощью флуоресцентных меток, подсвечивающих известные последовательности ДНК этих микробов. Локиархеи продемонстрировали разнообразие размеров и форм: яички, палочки, прутики, ниточки – многоликий хитрец Локи одобряет. Хеймдалльархеи же оказались единообразными, но чрезвычайно приятными на вид кругляшами, в центре которых даже можно разглядеть скопление ДНК. Для прокариот последняя черта весьма необычна, а вот у эукариот подобное скопление интерпретировалось бы однозначно: ядро!
(Обновление. Буквально на следующий день после публикации этого номера стало известно о японском исследовании, в ходе которого удалось не только получить первые четкие снимки локиархей, но и впервые вырастить их популяцию, изучить и описать как вид! Подробнее об этом читайте в нашей отдельной статье.)
Ядерный вопрос
Вопрос в том, на каком этапе асгардархея стала истинной эукариотой. Слово «эукариота» (от греч. εύ – «хорошо» и κάρυον – «ядро») означает, что клетка содержит хорошо оформленное ядро. Но вот на какой стадии магистрального развития прокариоты обзавелись ядром, а главное – как это случилось, до сих пор является загадкой.
Наиболее простой способ создать ядро – вобрать в себя часть клеточной мембраны и отпочковать вместе с ДНК, оказавшейся внутри пузырика. Некоторые прокариоты способны формировать компартменты – например, бактерии из группы Planctomycetes, обладатели сложной системы внутренних мембран, у которых даже есть нуклеоид, окружающий ДНК. Более того, на некоторых внутренних мембранах планктомицета Gemmata obscuriglobis обнаружены поры, удивительным образом похожие по структуре на ядерные поры эукариот, только меньше по размеру и иного аминокислотного состава. Поразительный пример конвергенции! Почему бы и предкам эукариот не научиться мастерству компартментализации?
Константин Мережковский сто лет назад полагал, что ядро, как и хлоропласты, появилось в результате эндосимбиоза. Сегодня определенной популярностью пользуется гипотеза, согласно которой ядро – это бывший гигантский вирус, однажды проникший в архею. Некоторые ученые считают, что ядро – это остаток археи, случайно оказавшейся внутри бактерии, а цитоплазма со всеми мембранами у нас бактериального происхождения (затем эта химера включила в себя предка митохондрий, то есть эукариотная клетка – это тройственный симбиоз двух бактерий и одной археи). Еще одна гипотеза гласит, что архея выпускала за свои пределы ложноножки для обмена веществами с приютившимися рядышком бактериями, а потом эти ложноножки попросту слились друг с другом в цитоплазму, тело археи осталось ядром, а бактерии превратились в митохондрии:

В пользу этой гипотезы говорит то обстоятельство, что внутри эндоплазматической сети (но не на поверхности клеток эукариот) имеются белки, похожие на остатки архейной клеточной стенки. Гипотеза не противоречит тому факту, что биохимически ядерные мембраны эукариот – бактериального типа: гены архей, ответственные за синтез мембраны, могли в ходе эволюции уступить бактериальным генам, полученным в результате обильного горизонтального переноса. Недавно в Черном море выловили бактерию, у которой есть гены для построения как бактериальных, так и архейных липидов. Наши предки археи, пошедшие по пути эукариотизации, в какой-то период тоже могли обладать мембранами смешанного типа.
Как бы там ни было, ядро, при всей важности его содержимого, всего лишь пузырек, облаченный в двойную мембрану, которая является частью системы внутриклеточных мембран – эндоплазматической сети. Может ли микроб, который обладает такой сетью и ядром, считаться уже готовой эукариотой? Ядерные мембраны, несомненно, создали условия для усложнения генома и механизма его работы, одновременно оберегая ДНК от проникновения в нее чужеродного генетического материала. Но какой толк прокачивать геном, если для обеспечения его работы неоткуда брать лишнюю энергию? Даже если бы эволюция потворствовала ядерной инновации, организмы едва ли ушли бы в своем развитии дальше планктомицетов или асгардархей, не обзаведись они источником необходимой энергии.
Таким источником стали митохондрии – одомашненные протеобактерии. Они были среди тех, кого археи обняли ложноножками и приютили в своей клетке, согласно вышеописанной гипотезе. По мнению первооткрывателей асгардархей, предки эукариот привлекали протеобактерий излишками водорода и электронов, которые оставались после окисления органических молекул (например, жирных кислот и бутана), а потом, когда связь между археями и бактериями стала совсем уж интимной, выяснилось, что и от последних первым есть прок! Будущие митохондрии умели дышать кислородом и оказались настолько энергетически выгодным приобретением, что некоторые специалисты прямо говорят: момент их включения в архейные клетки и момент возникновения эукариот – это одно и то же событие.
Слава митохондрии!
В свое время ученые были разделены на два непримиримых лагеря. Одни склонялись к тому, что прокариоты эволюционировали в эукариот постепенно, увеличиваясь в размерах и приобретая инновации одну за другой: ядро, цитоскелет, фагоцитоз (необязательно в таком порядке) и, наконец, митохондрии – как очередной шаг классической дарвиновской эволюции. Линн Маргулис была убеждена в таком развитии событий до самого конца. Другие считали, что эукариоты возникли в результате своего рода «большого взрыва», спровоцированного обретением митохондрий, – только после этого клетки начали расти и приобретать все характерные для эукариот инновации, включая ядро. Академические дебаты были столь горячи, что приверженцы разных точек зрения зачастую демонстративно уходили с выступлений друг друга на конференциях, громко звеня митохондриями.
Открытие асгардархей склонило чашу весов в сторону первых: эти прокариоты уже, по-видимому, обладают близкими аналогами или даже зачатками некоторых эукариотических признаков. Кроме того, биоинформатические исследования белков эукариот, унаследованных от прокариот, показывают, что происхождение самых древних из них связано с археями (ядерные белки и выполняющие функции репликации, транскрипции и трансляции ДНК), вторые по древности – белки бактериального происхождения, полученные с помощью горизонтального переноса генов (функции этих белков связаны с мембранными структурами – эндоплазматической сетью, аппаратом Гольджи), ну а белки протеобактерий были приобретены эукариотами позже остальных. Иными словами, митохондрии вошли в состав клетки, уже порядком эукариотизовавшейся. Тем не менее именно их включение обеспечило ее потомкам эволюционный успех.
Без митохондрий у микробов и шансов не было стать полноценными эукариотами. Чтобы усложняться, нужен крупный геном. Эукариотические геномы в десять, а то и в десять тысяч раз больше прокариотических (хотя, например, у почвенных бактерий Myxococcus xanthus геном побольше иных эукариотных паразитов, таких как микроспоридии Encephalitozoon cuniculi). Но чтобы копировать огромный геном, а тем более производить белки на основе генетической информации, необходимо колоссальное количество энергии. На синтез белков может уходить до 80% энергии всей клетки! Если прокариота мечтает увеличить свой геном в десять раз, то ей стоит озаботиться вопросом, где взять в десять раз больше энергии на новые белки.
Она может увеличиться сама, увеличив и общую поверхность мембран, через которые осуществляются все процессы, связанные с производством энергии, – прежде всего перекачка аденозинтрифосфата (АТФ), универсального носителя энергии в живых клетках. Но в таком случае возрастет и количество белков бактерии, на чей синтез будет тратиться больше энергии, чем зарабатываться. Прокариоты заключены в энергетические кандалы, которые не позволяют им стать большими и сложными. Лишь один раз им удалось вырваться из этого плена – после того, как они обрели митохондрии.
Внутри каждой из митохондрий полно мембранных складочек – а значит, и поверхностей для выработки энергии. Еще у них есть свой собственный набор из 13 генов для регуляции этих процессов – независимый от центрального генетического аппарата эукариотной клетки, что находится в ядре. Так что никакой бюрократии и проволочек. Митохондрии – автономные энергетические станции, не очень мощные поодиночке, но зато способные делиться и формировать целые команды «энергетиков», чтобы решать возникающие внутри клетки проблемы энергоснабжения.
У прокариот таких энергостанций нет – они сами по себе энергостанции. Некоторые даже способны складывать внутрь себя мембрану с целью увеличения поверхности для производства энергии. Но у них нет дополнительных ДНК-администраций, готовых взять на себя функции управления внутриклеточной энергетикой, чтобы разгрузить центральную ДНК и тем самым предоставить ей возможность осуществлять всякие эволюционные эксперименты. Вступив в союз с бактериями, археи делегировали им энергопроизводство – и отныне могли позволить себе расширять геном, увеличиваться в размерах, нарабатывать инновации, одомашнивать других микробов, эволюционировать в доселе немыслимых направлениях.
Сливы митохондрии
Казалось бы, столь важная роль митохондрий в жизнедеятельности эукариот подразумевает, что у всех эукариот они должны быть непременно. Как бы не так! Взять, к примеру, лямблий (Giardia) – одноклеточных жгутиконосцев из супергруппы Excavata, обожающих заражать кишки и вызывать диарею. Это явные эукариоты – у них даже не одно ядро, а два! Но митохондрий – нетути.
И таких протистов без митохондрий не меньше тысячи, по большей части это паразиты. Одно время их рассматривали как реликтовых потомков примитивных эукариот, не успевших обзавестись митохондриями, был даже соблазн выделить их в отдельное царство Archezoa. Однако впоследствии у них были выявлены гены протеобактериального происхождения – то есть митохондриями их предки все-таки обладали, просто утратили их за ненадобностью в условиях бескислородного проживания, хотя часть их генов и осталась, перекочевав в ядро. У некоторых, включая лямблий, даже обнаружились нефункциональные рудименты митохондрий – митосомы, у других эукариот имеются гидрогеносомы, которые тоже считаются деградировавшими митохондриями.
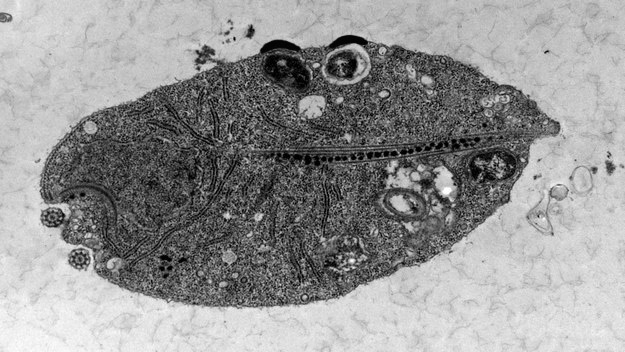
А вот у жгутиконосца Monocercomonoides, тоже из экскават, не то что митосом, а даже митохондриальных генов не осталось – это первый известный науке эукариотический организм без каких-либо следов митохондрий. Энергетический обмен он осуществляет за счет генов, приобретенных горизонтальным переносом от бактерий, не связанных родством с митохондриями. Всем своим видом этот протист демонстрирует, что эукариоты жить без митохондрий еще как могут. Вместе с тем никаким «архезоем» он не является – его положение на филогенетическом древе однозначно показывает, что он не примитивный, первично безмитохондриальный простачок, а специализированный, вторично утративший митохондрии дегенерат.
Есть и такие чудаки, у которых митохондрия есть, а генов в ней нет. Такова паразитическая морская динофлагеллята Amoebophyra ceratii, заражающая других, фотосинтетических динофлагеллят, что устраивают цветения в океане. Она лишилась пластид (органелл для фотосинтеза) вместе с их генами, зато у нее остались две выраженные митохондрии – только в них нет никакой митохондриальной ДНК. Тем не менее они прекрасно справляются со своей функцией – дышат кислородом, генерируют энергию. По всей видимости, происходит это за счет генов, некогда переданных ими в высшую инстанцию – ядро. Это первый известный организм, чьи митохондрии работают без собственной ДНК. Остальные эукариоты не настолько одомашнили свои митохондрии, как эта бестия.
Линн Маргулис верила, что когда-нибудь биологи отыщут наконец архезойную эукариоту – «архепротиста», у предков которого никогда не было митохондрий, – достанут его из какой-нибудь экологической тмутаракани, где он миллиарды лет успешно скрывается от кислорода и науки, свято храня тайны происхождения эукариот. Но пока что все известные случаи отсутствия митохондрий у эукариот признаны вторичными. Скорее всего, у общего предка всех ныне живущих эукариот митохондрии были. Это не означает, естественно, что более ранние, базальные эукариоты ими обладали. Никто до сих пор не знает точно, когда состоялся счастливый эндосимбиоз – до возникновения ядра или же после.
Об уникальности первичных эндосимбиозов
Один раз попробовали – и понеслась. Эукариоты по жизни оказались открыты для всяческих эндосимбиозов. Примерно миллиард лет назад кто-то из них проглотил цианобактерию, но не переварил, и она стала фотосинтезировать ему во благо, размножаться внутри него и вместе с ним. Так появились растения, или, по-умному, архепластиды (супергруппа Archaeplastida). Со временем потомки той цианобактерии превратились в пластиды – муропласты (цианеллы) глаукофитов, родопласты красных водорослей и хлоропласты зеленых водорослей и высших растений. Затем одноклеточных красных и зеленых водорослей стали поедать и периодически одомашнивать внутри себя разные протисты, а вот с глаукофитами не сошлись характерами – по крайней мере вторичные пластиды глаукофитового происхождения науке не известны. Зато известны третичные пластиды – потомки протистов с вторичными хлоропластами, прирученных другими протистами. Эволюция явно не скупилась на пластидные эндосимбиозы эукариот. Однако первичный – с бактерией – был всего один.

Первичные эндосимбиозы, в которых бактерия становится клеточной органеллой, вообще большая редкость. За всю историю жизни их можно пересчитать по пальцам руки трехпалого ленивца: (1) митохондрия эукариотной клетки, (2) пластида растительной клетки, (3) хроматофор паулинеллы. Три вида раковинных амеб Paulinella (относятся к супергруппе Rhizaria) фотосинтезируют благодаря хроматофору (их может быть два) – особой сарделькообразной пластиде, произошедшей от некой цианобактерии, захваченной не так уж и давно, примерно 100 млн лет назад, в мезозое. Как и в случае митохондрий с хлоропластами, часть генов хроматофора перекочевала в ядро хозяйской клетки (хоть и не столь значительная), и поэтому он не может выжить отдельно от паулинеллы, то есть является типичной органеллой (хоть и относительно юной в эволюционном смысле).
Почему первичные эндосимбиозы столь редки? Возможно, бактерии, потенциально способные принести пользу хозяину, обычно не приспособлены к внутриклеточной жизни и не могут долго выживать в хозяйской цитоплазме. А вот некоторые паразитические бактерии даже могут в ней размножаться – например, альфа-протеобактерии Rickettsia, родственницы предков митохондрий. Есть даже гипотеза, что предок митохондрий тоже был паразитом: он умел противостоять «иммунитету» архей, а значит, был предрасположен к эндосимбиозу – оставалось лишь перепрофилироваться из энергетического паразита в поставщика энергии путем случайной смены полярности белка, качающего АТФ через бактериальную мембрану. Предковая для пластид цианобактерия вряд ли была паразитом, но обосноваться в клетке-хозяйке ей могла помочь паразитическая бактерия из группы хламидий, проникшая туда вместе с ней, – в ядерных геномах растений сохранились гены хламидиального происхождения. Каким образом другая цианобактерия прижилась в паулинелле, пока непонятно. Но не стоит недооценивать созидательную мощь паразитизма!
Конечно, ни о каких растениях с паулинеллами и прочими протистами не пришлось бы и мечтать, не случись 1,5–2 млрд лет назад союза асгардархей и протеобактерий – самого первичного из первичных эндосимбиозов. Вот уж поистине уникальное событие! Вдумайтесь: за 3,5 млрд лет существования прокариот они смогли вступить в перспективный союз всего лишь раз, несмотря на то что постоянно контактируют друг с другом там и сям. Может, были и тысячи других попыток, но успехом они не увенчались – альтернативные формы эндосимбиотической жизни, отличной от эукариотической, на нашей планете неизвестны.
Ничтожная вероятность подобных событий может объяснить, почему нам до сих пор не встретилась хоть какая-то мало-мальски разумная жизнь за пределами Солнечной системы. В нашей галактике Млечный Путь миллиарды планет, многие из них в зоне обитаемости, и наверняка есть такие, где химические условия позволили зародиться живому. Однако без случайного и судьбоносного симбиоза любая подобная жизнь обречена оставаться микробной. Микробов во Вселенной много, но сложная, тем более разумная жизнь невероятно редка. И если все-таки разумные инопланетяне существуют – у них должны быть аналоги митохондрий.
Текст: Виктор Ковылин. Иллюстрация на обложке: Антонина Оснос
Все права на текст принадлежат нашему журналу. Убедительная просьба не копировать его в соцсети или куда-либо еще без договоренности с редакцией. Если хотите поделиться информацией с вашими подписчиками, можно использовать фрагмент и поставить активную ссылку на этот номер – мы будем рады. И конечно, будем очень признательны за любую поддержку нашего проекта. С уважением, Батрахоспермум.
Вас также могут заинтересовать статьи:
Древнейший родственник людей и водорослей объявился в Индии
Эндосимбиотическая матрешка червеца демонстрирует движуху
Как стать хорошим микробом-симбионтом